by Elizabeth H. Beers, Jay F. Brunner, and Bruce A. Barrett, originally published 1993; revised December 2007
Phyllonorycter elmaella Doganlar & Mutuura, (Lepidoptera: Gracillariidae)
Western tentiform leafminer has been a pest in Washington orchards since about 1980. It is a somewhat sporadic pest in most areas, with serious infestations occurring only about one to two years out of five. The frequency and intensity of leafminer outbreaks has been much lower since the early 2000s, likely because of shifts in pesticide programs for key pests. Problems are generally reduced where selective pesticide programs (those programs that have limited effects on non-pest and non-target insects) are used. This is an indirect pest, causing damage only to the foliage and not directly damaging or affecting the fruit. These two factors make it an ideal candidate for biological control.
There is some question as to western tentiform leafminer’s status as a different species from a closely related species, the spotted tentiform leafminer (Phyllonorycter blancardella) occurring the eastern and central US. In any case, the life cycle and damage of the two species (or races) are virtually identical.
Hosts
Western tentiform leafminer attacks primarily apple and crabapple; if the latter are used as pollenizers they may be heavily infested. It can attack pear, although this has been quite uncommon in Washington. It also attacks sweet and tart cherry, and has been locally severe on the former crop.
Life stages
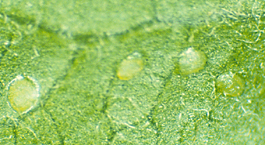
Eggs
Western tentiform leafminer eggs are small (0.3 mm in diameter), elliptical, and creamy to transparent. The transparency often makes them appear the same color as the leaf. Eggs are laid singly on the undersides of leaves and, depending on temperature, hatch in 5 to 16 days.
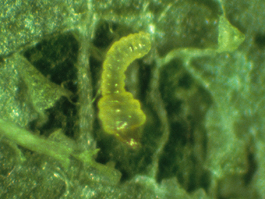
Larva
Western tentiform leafminer larvae have five instars (stages), and their entire life is spent inside the mine, protected from the exterior environment. The first three instars are relatively small and are referred to as the sapfeeders because they feed on the sap from the spongy mesophyll layer of the leaves. The sapfeeders are flattened and legless, with an enlarged segment just behind the head, giving them a “broad-shouldered” appearance. In the process of feeding, they separate the lower leaf surface from the tissue above. During this stage, the mines are small, only visible from the lower leaf surface, and appear as light green lines or blotches. The fourth and fifth instars feed more on the leaf tissues and are referred to as the tissuefeeders. These larvae have a more typical lepidopteran larval appearance, with a cylindrical body, three pairs of legs on the thorax. The change in feeding habit and the increased size of the larvae make the mine visible from the upper as well as the lower leaf surfaces. It also gives the mines a tent-like appearance from the upper leaf surface, with small white spots where the green tissue has been removed. The lower surface will often develop a “seam” during this stage, where the tissue is bunched together longitudinally down the center of the mine. When full grown, the larvae are about 4 mm long, cylindrical, and white to pale green. Before they pupate, the larvae turn yellow. Larval development takes about 24 days to complete for the spring generation, but occurs faster during the warmer temperatures during summer.

Pupa
The pupae are 3 to 4 mm long and change from bright tan when first formed to dark brown. The pupal period lasts about 10 days for the first three generations and extends through the winter for the fourth generation. The pupa pushes out through the end of the mine, and the adult emerges from the pupal case, leaving it behind partially lodged in the mine. The pupal skin remains attached to the leaf for a time after the adult has emerged.
Adult
Adult moths are slender brown moths with distinct silver to cream colored bands on the upper wings Their length (2.4 to 4.0 mm) varies between sexes and generations. The moths that emerge in the early spring from overwintering pupae tend to be larger and darker than adults of the other generations. Mating and egg-laying occur in the evening. A female will lay an average of 25 eggs.


Life history
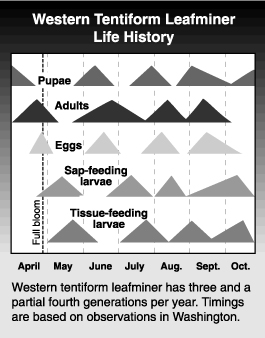 Western tentiform leafminer has 3 to 4 generations per year in Washington. Adults emerge from the overwintering pupal stage very early in the spring, about the same time green tissue is seen on apple buds. These adults lay eggs on the first unfolding cluster leaves. Eggs begin hatching just prior to bloom of apple. The eggs are laid on the underside of the leaves, and the first mines may be found shortly after bloom. The newly hatched larva moves directly from the egg into the leaf, and is never exposed on the leaf surface. The larvae feed on leaf tissue between the upper and lower leaf surface creating a ‘mine’. It feeds inside and enlarges the mine as it completes its development. The larvae pass through five instars (stages between molts), with the first three known as “sapfeeders”, and the final two called “tissuefeeders” (see description of stages below). The larvae pupate inside the mines, emerging as adult moths by cutting a slit in the end of the mine. Often the empty pupal case can be seen partially protruding from the mine. Adult emergence of the first generation begins about mid-June depending on the year and the region. Because only leaves associated with the flower cluster and very early shoot leaves are available to the overwintering adults for oviposition, first generation mines are confined to this area. The tree canopy is almost fully developed by the time the first generation adults emerge, thus this and subsequent generations lay eggs and produce mines throughout the tree. Eggs of the second generation are laid beginning in mid- to late June, and hatch in about 7 to 10 days. Mines develop more quickly in the warmer summer temperatures and emergence of the third generation adult begins in late July. The mines from this generation are found in August and early September. Depending on how warm the year has been, some of the pupae of the third generation may emerge as adults and begin a final (fourth) generation before freezing temperatures stop insect development. At the same time, some proportion of the third generation pupae will remain in the mines, fall to the ground with the leaves, and overwinter in this stage. Larvae that do not make it to the pupal stage before cold weathers sets in permanently do not survive the winter. The entire life cycle (egg to adult) requires 35 to 55 days, with shorter development times occurring in mid-summer, and longer times in cooler spring and fall weather.
Western tentiform leafminer has 3 to 4 generations per year in Washington. Adults emerge from the overwintering pupal stage very early in the spring, about the same time green tissue is seen on apple buds. These adults lay eggs on the first unfolding cluster leaves. Eggs begin hatching just prior to bloom of apple. The eggs are laid on the underside of the leaves, and the first mines may be found shortly after bloom. The newly hatched larva moves directly from the egg into the leaf, and is never exposed on the leaf surface. The larvae feed on leaf tissue between the upper and lower leaf surface creating a ‘mine’. It feeds inside and enlarges the mine as it completes its development. The larvae pass through five instars (stages between molts), with the first three known as “sapfeeders”, and the final two called “tissuefeeders” (see description of stages below). The larvae pupate inside the mines, emerging as adult moths by cutting a slit in the end of the mine. Often the empty pupal case can be seen partially protruding from the mine. Adult emergence of the first generation begins about mid-June depending on the year and the region. Because only leaves associated with the flower cluster and very early shoot leaves are available to the overwintering adults for oviposition, first generation mines are confined to this area. The tree canopy is almost fully developed by the time the first generation adults emerge, thus this and subsequent generations lay eggs and produce mines throughout the tree. Eggs of the second generation are laid beginning in mid- to late June, and hatch in about 7 to 10 days. Mines develop more quickly in the warmer summer temperatures and emergence of the third generation adult begins in late July. The mines from this generation are found in August and early September. Depending on how warm the year has been, some of the pupae of the third generation may emerge as adults and begin a final (fourth) generation before freezing temperatures stop insect development. At the same time, some proportion of the third generation pupae will remain in the mines, fall to the ground with the leaves, and overwinter in this stage. Larvae that do not make it to the pupal stage before cold weathers sets in permanently do not survive the winter. The entire life cycle (egg to adult) requires 35 to 55 days, with shorter development times occurring in mid-summer, and longer times in cooler spring and fall weather.
Damage

The leafminer’s impact on production and quality of fruit depends on the time of infestation, apple variety, tree vigor, number of mines, and additional stresses such as water stress or damage by other pests. No impact on fruit size or quality has been observed when populations average less than two tissue-feeding mines per leaf in the second generation. While two mines per leaf would not cause damage to apple in late June, populations this high could result in damaging numbers of mines in the third generation. When the third generation has occurred late, at the end of August or in September, no impact on fruit size or quality has been documented where there were fewer than five tissue-feeding mines per leaf. However, when the third generation has begun early, in late July or early August, densities of eight or more tissue-feeding mines per leaf have reduced fruit size and soluble solids in Red Delicious. The effect of the leafminer on fruit color has not been determined. In hot, dry summers when fruit is susceptible to sunburn, high leafminer infestations can aggravate this problem because infested leaves are curled or cupped, offering less shade.
Biological control
 At least four species of parasitoids are know to attack western tentiform leafminer in Washington, but one species of parasitic wasp (Pnigalio flavipes) is by far the most common. Other species include Sympiesis marylandensis, and two unidentified endoparasitoids. Pnigalio flavipes is an ectoparasitoid (the parasitoid larvae attach to and develop on the exterior of the leafminer larva), and thus are clearly visible during their life cycle. The adult female wasp searches leaves until she finds a mine. At that point, she paralyzes the leafminer larva using her ovipositor (the stinger, that doubles as an egg-laying tube) and may either kill the larvae and feed on its juices (called “host feeding”), or lay an egg near the leafminer larva. The choice of whether to feed or lay an egg depends partially on the leafminer’s body size; female wasps prefer more mature prey (tissuefeeders) for laying their eggs, and only feed on younger larvae (sapfeeder) for nourishment. In either case the leafminer larva is killed. Once the parasitoid egg hatches, the parasitoid larvae finds its way to the leafminer, attaches via its mouthparts, and begins sucking body fluids from the leafminer. By the time the parasitoid pupates, the leafminer larva is a shriveled husk. Pnigalio flavipescan attack all of the leafminer generations, although they usually increase to their highest density in the third generation. High levels of parasitism (>90%) can greatly reduce the overwintering leafminer population, and even modest levels (>30%) during the first or second generation may be all that is required to keep leafminer densities below levels such that no chemical control is necessary.
At least four species of parasitoids are know to attack western tentiform leafminer in Washington, but one species of parasitic wasp (Pnigalio flavipes) is by far the most common. Other species include Sympiesis marylandensis, and two unidentified endoparasitoids. Pnigalio flavipes is an ectoparasitoid (the parasitoid larvae attach to and develop on the exterior of the leafminer larva), and thus are clearly visible during their life cycle. The adult female wasp searches leaves until she finds a mine. At that point, she paralyzes the leafminer larva using her ovipositor (the stinger, that doubles as an egg-laying tube) and may either kill the larvae and feed on its juices (called “host feeding”), or lay an egg near the leafminer larva. The choice of whether to feed or lay an egg depends partially on the leafminer’s body size; female wasps prefer more mature prey (tissuefeeders) for laying their eggs, and only feed on younger larvae (sapfeeder) for nourishment. In either case the leafminer larva is killed. Once the parasitoid egg hatches, the parasitoid larvae finds its way to the leafminer, attaches via its mouthparts, and begins sucking body fluids from the leafminer. By the time the parasitoid pupates, the leafminer larva is a shriveled husk. Pnigalio flavipescan attack all of the leafminer generations, although they usually increase to their highest density in the third generation. High levels of parasitism (>90%) can greatly reduce the overwintering leafminer population, and even modest levels (>30%) during the first or second generation may be all that is required to keep leafminer densities below levels such that no chemical control is necessary.
Monitoring
Western tentiform leafminer can become a problem in almost any orchard, but particularly in those where it has recently become established. However, a high population one year does not mean it will be a problem the following year. A good sampling program and knowledge of treatment thresholds are keys to successful management of the leafminer.
As there is no known relationship between adults seen or trapped and the subsequent number of mines, sampling should be based on the number of mines. Though tissue feeding mines are the most obvious indication of a leafminer infestation, it is more important to sample sap-feeding mines to determine the need for treatment. Sap-feeding mines can be seen by picking the leaf, holding it toward the light and examining the undersurface.
1st generation
It is not usually necessary to sample mines in the first generation except to assess the amount of parasitism. There are generally few mines as only a few of the earliest leaves were exposed to attack. The potential impact on tree vigor and fruit size or quality from mines in these leaves is low.
However, eggs of the first generation can be sampled at pre-pink or pink to determine the potential number of mines per leaf. Take at least 50 leaves (5 from each of 10 trees) and, using a hand lens or binocular microscope, examine the lower surface for eggs. Because the number of leaves on the tree increases rapidly during this period, it takes three eggs per leaf at pink to average one mine per leaf at the end of the first generation.
To determine the impact of parasitism, select 100 leaves from 25 trees that contain tissue feeding mines in late May or early June. Tear open each mine and determine if the leafminer larva or pupa is alive. If the leafminer is dead, determine if death was due to parasite attack by looking for a parasite egg, larva or pupa. If the leafminer larva seems normal but is not moving, it has probably been stung by a parasite but no egg was laid. This should be counted as death due to parasite attack.
Calculate the percentage of parasitized leafminers by adding up the number of mines affected by the parasite, dividing it by the total number of mines sampled and multiplying by 100. If more than 35% of the mines are parasitized, the potential for biological control is good. However, it may still be necessary to monitor parasitism levels in subsequent generations.
2nd generation
A binomial sampling plan can be used for the second generation. This is based on a tally threshold of 2 or more mines per leaf. A total of 10 trees must be sampled to provide an accurate estimate of the population orchard-wide.
During late June and early July sample 20 leaves from the basal to mid-portion of a vegetative shoot and examine to determine how many contain 2 or more sap-feeding mines. Repeat this procedure for 10 trees in the orchard. The number of leaves over the tally threshold can be looked up in the sampling table to get the average mines per leaf. After values have been obtained for 10 trees, average the mines per leaf to determine the leafminer density for the orchard. In addition to the binomial sample, the second generation can be sampled using the same procedure as for the third generation. Pick a mid-shoot leaf on 5 shoots per tree, 20 trees per block, or 100 leaves.

3rd generation
If the third generation occurs early, in late July or early August, sample sap feeding mines in early to mid-August. The same sampling scheme used in the second generation will not work for higher populations of leafminer. To determine the density of mines in the third generation, pick a mid-shoot leaf from 5 shoots and count the total number of sap feeding mines. Repeat this sample of 20 trees (100 leaves total), accumulating the number of mines as you go. Divide the total number of mines by 100 to get the number of mines per leaf. If the second generation was treated or if sap-feeders do not appear until late August or September, it should not be necessary to sample the third generation.
Treatment thresholds
The following treatment thresholds for the western tentiform leafminer are based on the limited experience with this pest in the western United States and from recommendations used in the eastern United States for controlling its close relative, the spotted tentiform leafminer.
1st generation
The threshold of one sap feeder mine per leaf is seldom, if ever, exceeded. Pre-bloom treatments are suggested only if extremely high number of moths are seen at tight cluster to pink stage and eggs average more than three per leaf.
2nd generation
Treatments should be applied if sap feeder mines exceed two per leaf, especially if first-generation parasitism was low, less than 35%. If the second generation develops early, new sap feeder larvae are present in early June, and parasitism in the first generation was low, the threshold for treatment should be reduced to one mine per leaf.
3rd generation
If second generation treatments were applied, or if the third generation develops late, in late August or early September, no treatments should be required. However, if the third generation develops early with new sap feeders present in late July or early August, treatments should be applied if sap-feeder mines exceed five per leaf.
Management
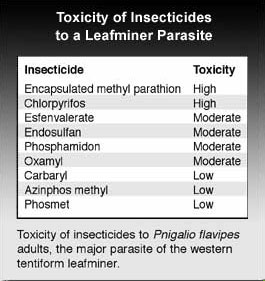
If biological control fails to suppress leafminer populations, chemical control may be warranted. A number of insecticides used in orchards may be toxic to leafminer parasitoids, (see above), and should be avoided if possible.
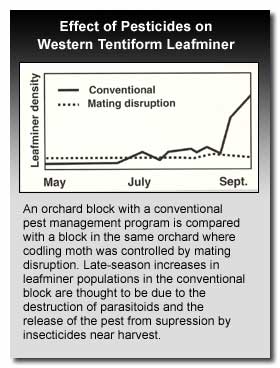 Treatment thresholds provide guidelines for making management decisions that will prevent damage by leafminers. However, because of the variability in cultivar sensitivity, growing region, tree vigor, and influences of other foliar pests, it is difficult for one threshold to encompass all possible situations. Ideally, the population density of the current generation should be used in conjunction with the parasitism level of the previous generation to make the best decision of need for control. For example, by assessing parasitism during the late tissuefeeder/early pupal stage of the first generation, and combining this information with the density of the second generation sapfeeder mines a decision for control of the second generation can be made. These two assessments will be made about 3 weeks apart with the last occurring just prior to a decision on the need for applying a chemical control.
Treatment thresholds provide guidelines for making management decisions that will prevent damage by leafminers. However, because of the variability in cultivar sensitivity, growing region, tree vigor, and influences of other foliar pests, it is difficult for one threshold to encompass all possible situations. Ideally, the population density of the current generation should be used in conjunction with the parasitism level of the previous generation to make the best decision of need for control. For example, by assessing parasitism during the late tissuefeeder/early pupal stage of the first generation, and combining this information with the density of the second generation sapfeeder mines a decision for control of the second generation can be made. These two assessments will be made about 3 weeks apart with the last occurring just prior to a decision on the need for applying a chemical control.
Unless the population is exceptionally heavy, treatment of the first generation is usually not necessary. Because the canopy is undergoing rapid expansion in the early part of the growing season, and the number of leaves is increasing quickly, a population estimate of three eggs or sap-feeder mines per leaf around bloom (late April) will be diluted to less than one mine per leaf by the end of the generation, in late May or early June. Thus, when the population is evaluated around bloom, the treatment threshold is three eggs or sapfeeder mines/leaf. The second generation is the most frequent target for chemical control, because it provides the best opportunity to assess both the pest density and the potential for biological control. However, if parasitism of the first generation is <30%, and sapfeeder mines of the second generation exceed 2 mines/leaf, an insecticide treatment is usually warranted. The third generation is usually not targeted for control, because it occurs too late to have a negative impact on the crop. However, in warm years when a fourth generation is likely or in orchards with late-maturing cultivars such as Fuji and Braeburn, the third generation may be more damaging. The same principles apply for the third generation: assess parasitism late in the second generation, and leafminers during the sapfeeding stage of the third. In this case, if the leafminer population exceeds 5 sapfeeding mines/leaf (and parasitism is <30%) then treatment may be warranted.
The primary target of most chemical controls for leafminer used is the sapfeeder larvae. Timing with these products is critical to success. The period between evaluation of the leafminer population and the correct timing to achieve optimal control can be very short. Older larvae (tissuefeeders) are generally harder to kill, and pupae are unaffected by insecticides. Eggs and adults may be killed to some degree, depending on the material. Because the sapfeeders are protected inside the mine, insecticides that penetrate the leaf (translaminar activity) are most effective, and spray adjuvants that aid in penetration can help enhance control.
Leafminers probably became resistant to the organophosphate insecticides many years ago (although in some cases, toxicity to parasitoids persisted) so these materials are no longer suitable as controls. Some carbamate insecticides remain effective, however, carbamate insecticides tend to be highly toxic to predatory mites, and thus use of alternative materials is preferable. The macrocyclic lactones (fermentation products of soil microorganisms) are quite effective against leafminer and are also toxic to other pests (and some predators). A newer class of insecticides, the neonicotinyls (or chloronicotinyls), vary in their effectiveness for leafminer control, even though most are characterized by translaminar if not true systemic activity. When used for other pests, some insect growth regulators may have long-term suppressive effects on leafminers, but are generally not useful for control of a damaging population. Particle films have also been shown to suppress leafminer populations, but appear to interfere with parasitism. Petroleum oils may suppress oviposition or cause egg mortality, but probably require multiple applications during the egg laying period.
