Written by Matthew Whiting and Rolando Martin. Washington State University, Updated June 2021
Profitability in sweet cherry (Prunus avium L.) production will depend on growers’ ability to produce superlative fruit and maximize packout. A significant cause of cullage (and therefore lost revenue and higher packing/sorting expenses) in several important cultivars is polycarpy (i.e., multiple pistils, or simply ‘doubling’). An informal survey of Washington cherry packing houses found that doubling ranges from between 5% and 15%, depending on the year and cultivar. In the warmer growing districts of California, doubling can be even higher (J. Culbertson and J. Columbini, pers. commun.). Indeed, we hypothesize that doubling will become increasingly problematic as growers plant orchards in warmer climates, attempting to garner premium returns for early-season sweet cherries. Moreover, our own research has shown that doubling may be as severe as 50% in a susceptible cultivar like ‘Tieton’. Clearly, the potential losses from doubling are economically significant.
A search for published research into causes of doubling reveals how little work has been done to better understand this disorder. Research from Japan showed a clear role of high temperature during the previous season (i.e., while flower buds are developing) on incidence of doubling for ‘Satohnishiki’ (Beppu et al., 2001). Doubling was greater in potted trees exposed to 35C/25C day/night temperatures between June and September than in untreated trees. In addition, the phenomenon appears to be unrelated to tree water stress (Beppu and Kataoka, 1999). Very little published research has investigated efficacy of potential preventative strategies. Whole-tree shading (ca. 80% shade) during flower bud development of ‘Satohnishiki’ reduced doubling in the subsequent season to 24% from 47% in unshaded control trees (Beppu and Kataoka, 2000).
At Washington State University in Prosser, the sweet cherry physiology program has been investigating the causes of multiple pistils and pragmatic means for reducing this disorder in cultivars common to the U.S. sweet cherry industry (Whiting and Martin, 2008). We began in 2005 with funding from the Washington Tree Fruit Research Commission and have taken an integrated approach, researching three distinct but related components of pistil doubling:
- microscopic evaluation of floral bud initiation and organ differentiation
- critical temperatures and periods of susceptibility
- practical strategies for reducing doubling
(1) Microscopic evaluation of floral bud initiation and organ differentiation
This work was undertaken to determine the time of initiation of each of the floral organs for sweet cherry cultivars that exhibit a ca. 5-week range in fruit maturity: ‘Chelan’, ‘Tieton’, ‘Bing’, ‘Skeena’ and ‘Sweetheart’. All plant material was collected from a research orchard 5 km north of Prosser, Washington. Trees were rooted on ‘Gisela®5’ and trained to a Y-trellised architecture in N-S rows. We documented the progression of floral organ differentiation beginning shortly after initiation (mid May) through November, collecting bud samples every couple of weeks from spurs on two-year old wood.
We discovered that flower bud initiation occurs at roughly the same time for all five cultivars, the first buds were apparent in mid to late May. There was no clear relationship between fruit harvest timing and timing of flower bud initiation among cultivars. However, the progress of floral organ differentiation for each cultivar appeared to be similar to sequence in fruit maturity – the earliest maturing cultivar ‘Chelan’ was the furthest advanced in bud development and the latest maturing cultivar ‘Sweetheart’ was several weeks behind in development of floral organs. Each cultivar followed a similar pattern of organ development and differentiation. A pentagonal whorl of sepal primordia was the first apparent stage of organ differentiation, followed by petals and anthers. Pistil primordia were visible in ‘Chelan’ buds by early August whereas the pistil was not visible in ‘Sweetheart’ buds until about 3 weeks later. It appears now that doubling is induced prior to pistil formation. Therefore, the onset of pistil development might be indicative of the tree being no longer susceptible to doubling. This generally occurred about 7 to 9 weeks after harvest.
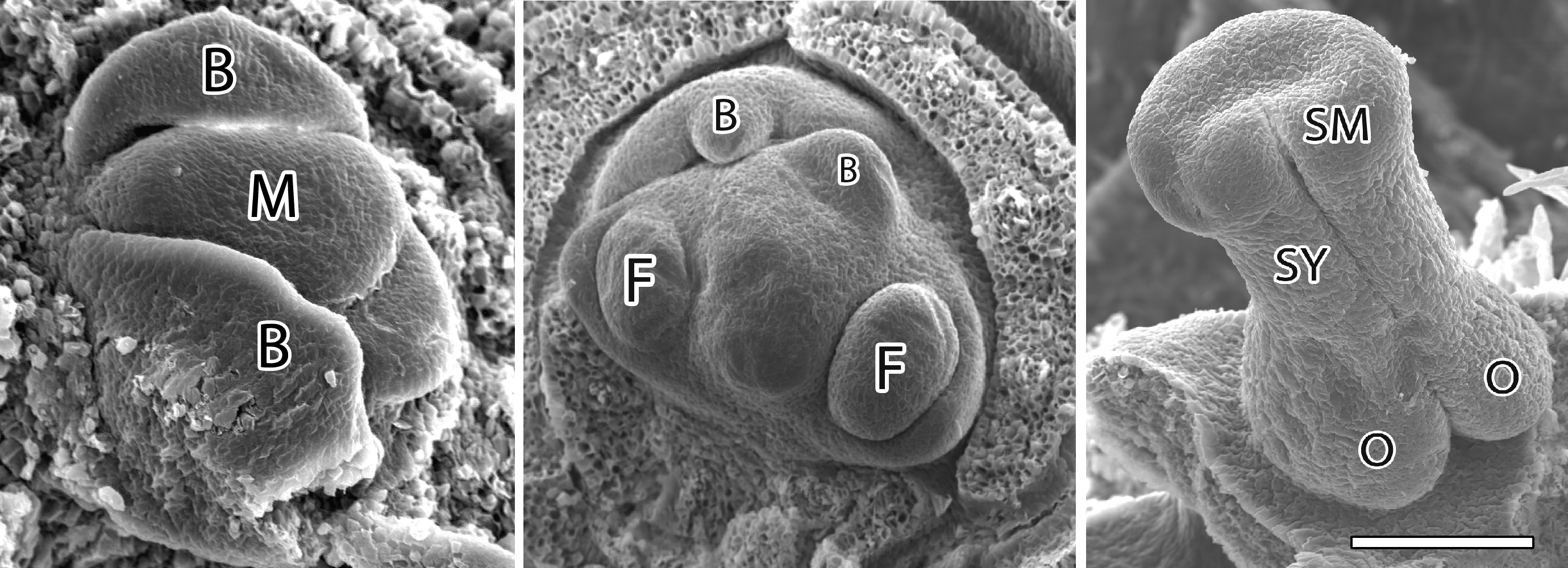
(2) Critical stages of organ differentiation and temperatures
We evaluated also the role of tissue temperature and seasonal variability in susceptibility to doubling by heating and cooling entire 2-year-old ‘Bing’ spurs in the orchard throughout key stages of floral organ differentiation. We continuously maintained a tissue temperature differential of ambient +9F for heated spurs and ambient -9F for cooled spurs throughout the treatment period (Figure 2). We applied heating/cooling treatments for 3 ca. two-week intervals in 2005 (early July to early August) and 6 similar intervals in 2006 (early July to early September). Doubling was then assessed by counting multiple pistils on treated spurs during bloom in the season subsequent to treatment.
We discovered that ‘Bing’ buds are susceptible to high temperature-induced doubling from late July to early September. Our data from 2005/2006 suggest that doubling susceptibility is highest in early August, when we were able to induce 15% doubling (data not shown). This is a stage of differentiation characterized by early sepal differentiation and no pistil formation. Our attempts to induce doubling in June and early July were unsuccessful, suggesting that buds are not susceptible at early stages of development (not surprising because our microscopy images reveal no floral differentiation at that time of year). In addition, once pistil growth progresses to the stage illustrated on the right in Figure 1, buds are no longer susceptible. Initial stages of pistil development appear to occur in late August and early September for ‘Bing’. However, we documented variability (1 to 2 weeks) in organ differentiation among individual flowers within a single reproductive bud (i.e., within a cultivar, some flowers are more/less advanced in development than others). Therefore, not all flowers in a bud may be at a susceptible/resistant stage of development. This may explain why doubling can be heterogeneous within a bud.
We discovered also that differentiating buds exposed to high temperatures prior to the susceptible period (i.e., late June, early July), acquire resistance to subsequent high temperatures during the susceptible stage of bud development. This discovery helps explain why the sweet cherry industries in Washington and Oregon observed fewer doubles in 2007 than expected – a week of temperatures with daily highs between 98F and 107F in late June ‘acclimated’ buds and therefore the high temperatures the region experienced in late July did not induce as much doubling as anticipated.

(3) Practical strategies for reducing doubling
The third part of this integrated research program investigated potential practical strategies for reducing doubling. This research was cooperative with industry and took place in commercial ‘Tieton’ orchards around the state. Our treatments compared the effect of over-tree evaporative cooling, Surround®, and in one orchard, a 20% shade. The treatments were initiated from mid July and terminated in mid August of 2006. We assessed doubling on representative limbs at bloom, and the same limbs and entire tree at harvest in 2007. In each orchard, three applications of Surround® were necessary to maintain coverage. Over-tree evaporative cooling was applied only when air temperatures exceeded 93ºF. Cooling was applied for twenty-minute intervals followed by ten minutes without cooling for as long as temperatures were high enough.
In a 5th-leaf ‘Tieton’/’Gisela®5’ orchard, natural doubling in 2007 was about 30%, on a whole-tree basis (Figure 3). This high incidence of doubling is not unusual for young ‘Tieton’ trees and reflects the potential severity of the disorder. However, each preventative treatment reduced doubling significantly, with over-tree evaporative cooling being the most effective. Shade, Surround®, and evaporative cooling reduced doubling by 37%, 45%, and 50%, respectively. By eliminating 50% of the doubles, an additional 2400 lbs of fruit per acre was salable (yield was 8 tons/acre). We found similar reductions in doubling from evaporative cooling and Surround® treatments in a mature ‘Bing’/Mazzard orchard. The Surround® treatment in 2006 reduced doubling in 2007 by 52% and cooling reduced doubling by 44%. Natural doubling in this ‘Bing’ orchard was only 4% however.
Over-tree evaporative cooling and Surround® show promise for reducing cullage from multiple pistils. Our data suggest that preventative treatments should be targeted for late July and August when temperatures are high (mid 90s).

In these trials we also measured canopy tissue temperature using thermocouples throughout entire canopies. We found the greatest reductions in tissue temperature with over-tree evaporative cooling (nearly 9F lower than untreated). Surround applications and a 20% shade treatment were moderately effective (ca. 3.5-4F reductions). Under-tree microsprinklers were ineffective for reducing canopy temperature, and even increased temperature slightly (ca. 2F). It is not recommended therefore, to run under-tree sprinklers in any attempt to reduce doubling. The increased humidity drives down the vapor pressure deficit, effectively reducing natural evaporative cooling from the canopy.
Contact

Matthew Whiting
mdwhiting@wsu.edu
Literature cited
Beppu, K, T. Ikdea, and I. Kataoka. 2001. Effect of high temperature exposure time during flower bud formation on the occurrence of double pistils in ‘Satohnishiki’ sweet cherry. Scientia Hortic. 87:77-84.
Beppu, K. and I. Kataoka. 1999. High temperature rather than drought stress is responsible for the occurrence of double pistils in ‘Satohnishiki’ sweet cherry. Scientia Hortic. 81:125-134
Beppu, K. and I. Kataoka. 2000. Artificial shading reduces the occurrence of double pistils in ‘Satohnishiki’ sweet cherry. Scientia Hortic. 83:241-247.
Whiting, M.D. and R. Martin. 2008. Reducing sweet cherry doubling. Good Fruit Grower. 59(4):24-26.
